

A big open question in 21st century science is how life began here on Earth. The metabolism-first scenario just might be the best one.
Planet Earth is overrun with life. Lakes, rivers, seas and oceans are teeming with it, from the surfaces all the way down to the bottom, often at depths of miles and miles. The land, both above and below ground, is packed with living organisms of varying size, mass, and complexity, including plants, animals, and fungi. Even the atmosphere houses a wide variety of life forms, from birds and insects to microbes found far above the highest mountain peaks. All told, more than 8 million species of organisms are currently represented on Earth, totaling over half a trillion tonnes of carbon in overall biomass.
We can trace our evolutionary history through time, with notable milestones including:
- the development of mammals and plants,
- the emergence of sexual reproduction and multicellularity,
- the biological creation of oxygen in our atmosphere, and
- the beginnings of photosynthesis.
We have fossil evidence of life existing 3.8 billion years ago, but the start of it all — the origin of life itself on Earth — remains an unsolved puzzle. Although many theories and scenarios abound, one of the least-talked-about may actually be the most likely: a metabolism-first scenario for life’s beginnings.
Here’s why recent research, only conducted in the last few years, may revolutionize the story of life’s emergence on our planet.
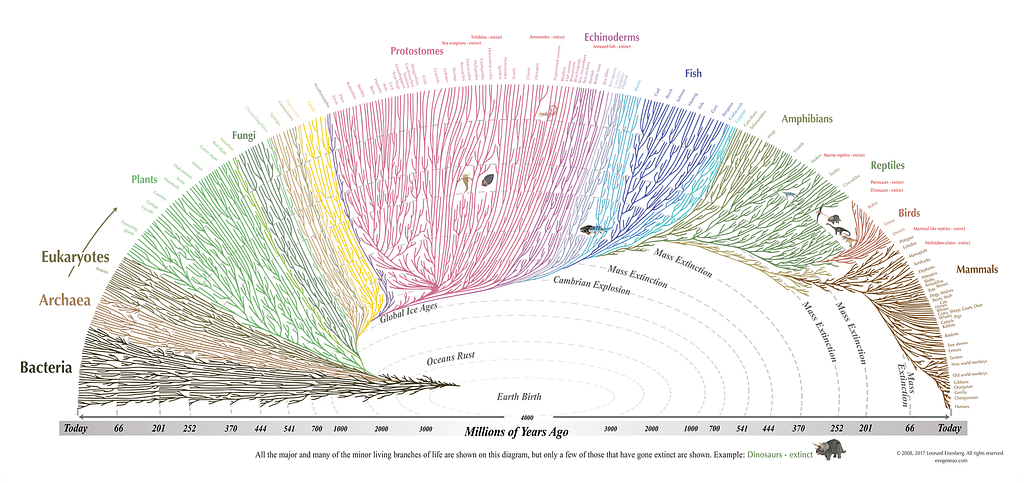
The 8+ billion species of organisms found on our world today possess an enormous diversity of properties. Some are large, some are small; some are complex, some are simple; some live only under specific, extreme conditions, while others thrive in a wide variety of terrestrial environments; some complete their life cycle in only a few hours, while others survive for decades, centuries, or even millennia. There doesn’t seem to be a universal set of conditions — at least, among these and many other common metrics — that you can apply to life.
And yet, there are at least five properties that are universal to all modern life-form:
- All forms of life collect resources of some type from their environment.
- They all possess a metabolism, where energy is extracted from an external source to achieve the organism’s metabolic goals.
- They all have the capacity to reproduce, making offspring that are partially or even wholly identical to the parent organism.
- They all have a separate “inside” and “outside,” delineated by the existence of a cell wall or cell membrane, creating a boundary between the organism and the external environment.
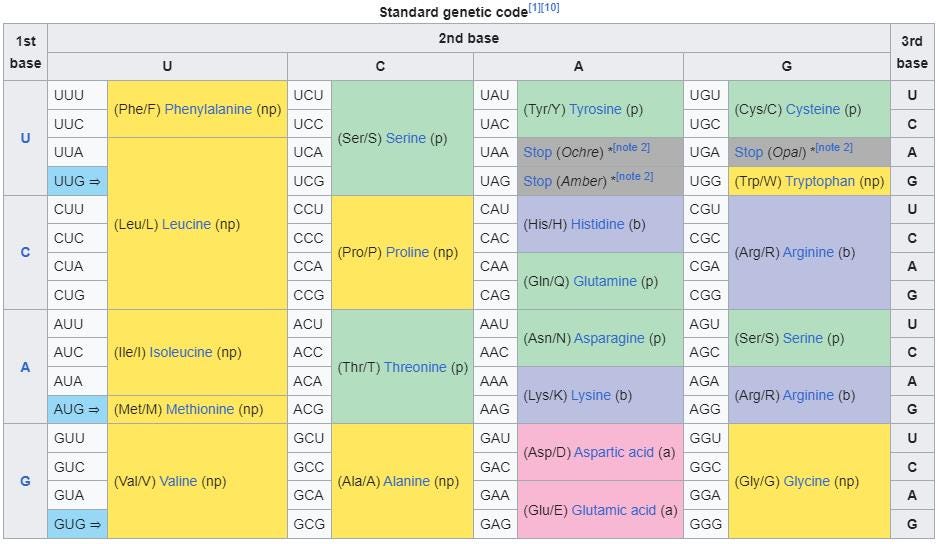
- And they all have, within them, some sort of genetic code that enables proteins to be synthesized and other life processes to occur.
It’s very unlikely that all of these properties arrived simultaneously, in a fully developed fashion. One of them must have come first.
Many look to the raw chemical ingredients at the heart of our understanding of life today, and advocate for their prominence — and even their primacy — in the origin of life. After all, they argue, those raw ingredients, including all of the nucleobases used in terrestrial life, are found extraterrestrially: in asteroids, comets, and other primordial bodies left over from our Solar System’s formation. Even though the odds of a string of those nucleobases randomly forming in a sensible order that encodes a successful protein are astronomically small, they still argue that it only takes one such success to lead to life, no matter how remote the odds.
Others instead point to the necessity of an inside-outside difference. They argue that the first life-forms required a protective layer to withstand the harshness of early Earth’s environment, complete with:
- solar radiation,
- lightning storms,
- heavy bombardment by objects from space,
- and copious volcanic activity,
to avoid proteins denaturing and their sensitive inner constituents from disassembling.
Those who advocate a cellular structure first often rely on the presence of lipids in an aqueous environment to support their argument. But the question of how the information to create such a structure could arise concurrent with all the other necessary functions that life would need to have remains; a “barrier-first” scenario won’t necessarily lead to life processes being carried out.

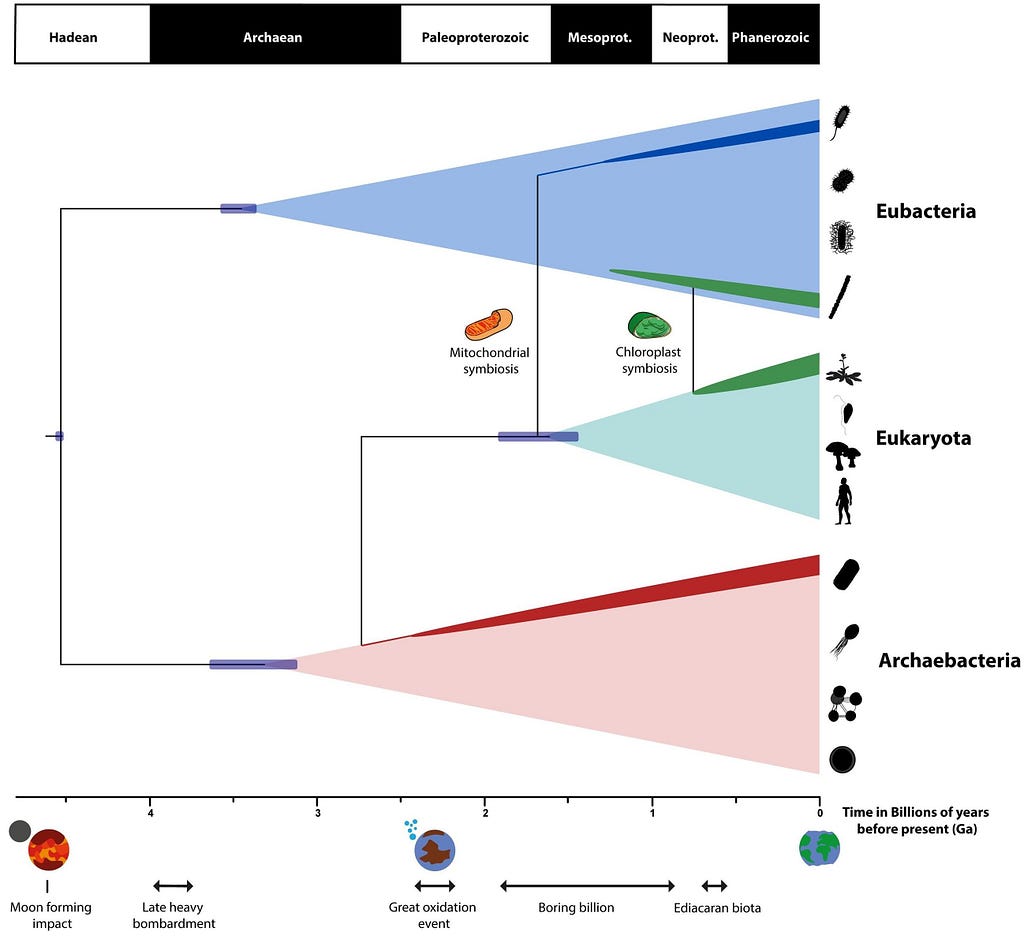
So how did life originate? Efforts to answer this question took a huge step forward in 2010, when a landmark paper integrated the evidence across the entirety of life on the planet with modern phylogenetics and rigorous probability theory. Previously unchallenged assumptions — such as the notion that similarity in genetic sequences necessarily implies genetic kinship, or that universal common ancestry was a requirement — were thrown out in favor of assumption-neutral tests. Horizontal gene transfer between barely-related species, including species in different kingdoms or phyla, was considered, along with fusion events. No stone was left unturned.
The results of conducting the formal test were as follows:
- Overwhelmingly, the idea that all extant life shares a universal common ancestor is favored, and all alternative hypotheses are disfavored.
- Horizontal gene transfer absolutely does occur, but it is extremely unlikely to occur between organisms descended from separate incidents of cell formation, as their genes would be converted into non-coding segments.
- The fact that the same 22 amino acids — and those 22, out of more than 80 known to naturally occur — are found in biologically produced protein molecules is additional strong chemical evidence in favor of universal common ancestry.
But even with this insight into life’s development and history on Earth, we still couldn’t draw definitive conclusions about its origins.
That’s why the newest approach isn’t to choose an assumption about what came first, but rather to begin with the conditions that must have been present on primordial Earth and work backwards: what types of reactions would have been likely to occur, or even inevitable?
In the beginning, what would become our Solar System was no more than an enriched cloud of primeval gas. Its composition was about 70% hydrogen, 28% helium, and around 1% oxygen, followed by smaller amounts of other elements, including carbon, neon, nitrogen, iron, silicon, sulfur, calcium, phosphorous, potassium, sodium, magnesium, and many others. Some of those atoms were bound up into molecules, including sugars, amino acids, nucleobases, aromatic molecules, and so on. Most of the mass is drawn to the center, where it will eventually form our Sun, but a substantial amount collapses into a rotating disk that surrounds the central protostar: a protoplanetary disk.
While the lightest elements in the inner part of the disk — hydrogen and helium, as well as light species of ice, such as nitrogen, methane, and carbon dioxide ices — boil and/or sublimate away, the heavier elements coalesce, forming longer-chain, more complex molecules. Over millions of years, imperfections arise in that protoplanetary disk, leading first to protoplanets and later, as the Solar System matures, full-fledged planets.

Early Earth was rife with violent events. The most famous is likely the collision with the protoplanet Theia about 4.5 billion years ago, which led to the formation of our Moon, with a subsequent period of heavy bombardment likely persisting for hundreds of millions of years thereafter. A combination of volcanic events and impacts from comets and asteroids led to the creation of oceans and an atmosphere, and precipitation on the planet’s early, highly uneven terrain led to the formation of freshwater stores, including rivers, lakes, and ices.
Although we colloquially use the phrase “boiling the ocean” to describe an overly ambitious, but practically impossible approach to problem solving, there’s a germ of a sound idea in that phrase that’s relevant. Since oceans are made of mostly water, but with many dissolved or suspended other particles and ions within it, “boiling” provides a method for removing the water while leaving the remaining contents behind. If you were to take even a large scoop of ocean water and began to boil it, you’d lower the fraction of water that’s present, removing it step-by-step, while leaving all of the dissolved and undissolved contents behind.
Now consider the various aqueous environments that our planet possessed early on, and you’ll see why the freshwater stores that formed over volcanically active areas — known as hydrothermal fields — are where Earth’s primordial ingredients were most concentrated. As their water evaporated, the density of organics within them — sugars, amino acids, nucleobases, ions, and much more — increased.

How can we be certain about the raw ingredients that were present? The best proxy for that is the composition of asteroids, comets, and meteorites. When we look inside these primitive objects — many of which we can date back to ~4.56 billion years ago — we find:
- they contain over 80 unique amino acids,
- many amino acids come in both left-handed and right-handed, even though all the ones that participate in life processes on Earth are exclusively left-handed,
- carbon-based organic molecules are also present, from the simple (like sugars) to the intermediate (like hexamethylenetetramine) to the complex (like polycyclic aromatic hydrocarbons),
- and, quite profoundly, all five of the nucleobases that encode genetic information on Earth are present in meteorites as well.
If you create a nutrient and organic-rich environment in the lab, you can do things like make an early-Earth analogue. You can apply energy to it, enable phase changes, and allow long-term chemical synthesis to occur. Large, complex molecules easily emerge, including full-fledged nucleotides, complex proteins, and enzymes. You’ll synthesize not just sugars but polysaccharides and even starches, as well as molecules bearing many similarities to modern cholesterols, alcohols, and lipids.

Lots of complex, long-chained molecules are going to form in an environment such as this. Amino acids will assemble, link up, and form proteins. Most of those proteins will be completely inactive; they won’t perform any biologically useful functions. However, if you replace the neutral atom at the end of one of those proteinss with an ion — particularly with a heavy element ion, such as magnesium — then that protein becomes an enzyme. Suddenly, your previously useless protein gains the ability to do things like:
- cleave molecules in two,
- catalyze energy-releasing chemical reactions,
- and turn previously “useless” molecules into a source of food and/or nutrients.
This isn’t a scientifically validated scenario depicting how life on Earth must have gotten its start, but rather a plausible scenario for how, before there was anything else (a cell membrane, a string of nucleic acids that encoded information, or even the ability to reproduce), there could have been molecules conducting metabolic activity.
As was first shown in a groundbreaking paper in 2013, this idea of a metabolism-first development of life should be considered, if not the default story of how biological life processes developed here on Earth, then at least a default component of the story.

This conversion from a useless protein to a useful enzyme can occur not only in hydrothermal field situations, but in tidepools, around hydrothermal vents at the bottom of the ocean, at the sea/air interface, or at other locations where non-equilibrium conditions persist. Amino acids interact and smack into one another, spontaneously forming and breaking bonds. Ions come along and bind to these primitive peptides, creating enzymes. Although these molecules are fragile and easy to destroy or denature, they’re very numerous and were found in high concentrations in these early environments, creating copious possibilities — set by the so-large-it’s-barely-fathomable mathematics of combinatorics — that truly boggle the mind.
Some of the proteins that formed likely gained the ability to perform specific functions merely by chance. These functions might have included the ability to:
- hoard resources, including specific peptides that can serve as food,
- split/recombine other molecules in a way that liberates usable energy in the process,
- “bite” or cleave other useful molecules, releasing energy while remaining intact themselves.
Whatever the case, the 2013 paper showed that the spontaneous creation of these metabolic peptides is all but inevitable. Then, less than 10 years ago, another incredible biological breakthrough was made into origin-of-life research: the discovery of RNA-peptide coevolution.
In an aqueous environment, nucleobases — the genetic “letters” of structures like RNA, DNA, or even PNA (peptide nucleic acids) — may line up along the various amino acids in a peptide chain. If each amino acid can pair up with its corresponding three-nucleobase codon, which can then “peel off” and draw additional amino acids onto that genetic strand, they can effectively reproduce, to a high degree of accuracy, the original peptide chain.
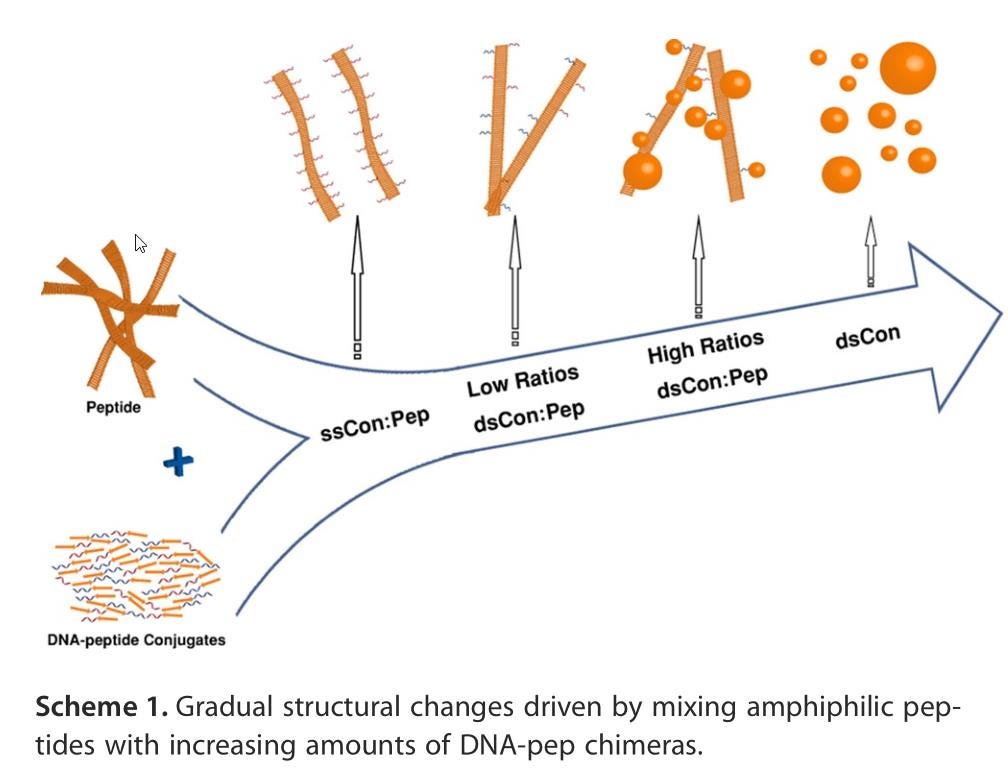
The RNA-peptide coevolution scenario, although new on the scene, has rapidly gained a following and is considered by many to be a leading theory for the origin of life not only here on Earth, but possibly anywhere the conditions for life’s emergence exist. The border between chemical and biological processes is blurry, but the idea of a primitive molecule that can metabolize nutrients found ubiquitously in its environment is highly attractive. If you then have an abundance of nucleic acids, and those nucleic acids can spontaneously align along the amino acid sequences that DNA, RNA, or even PNA can encode, you get a mechanism for another key component of life: replication.
If you have a metabolizing replicator that can successfully reproduce before its environment runs out of resources, denatures the molecule, or otherwise drives it to extinction, then the next steps can begin to fall into place, with the development of cell walls or membranes that delineate an “inside” of an organism from an “outside” chief among them.
We still have a long way to go in determining whether life is common, uncommon, rare, or even unique in the Universe; life here on Earth remains the only example we’re aware of. However, the clues to our origins aren’t just written Earth’s history, but also into the laws and conditions found throughout the Universe. If life happened here, it could happen elsewhere. Perhaps our first detection of life on other worlds will happen in the very near future. Perhaps that discovery, coupled with some key insights into how life first arose on Earth, will allow us to finally understand if metabolism, rather than cellular structure or an underlying genetic code, is the key that unlocks life’s emergence in general.
Starts With A Bang is written by Ethan Siegel, Ph.D., author of (affiliate links following) Beyond The Galaxy, Treknology, The Littlest Girl Goes Inside An Atom, and Infinite Cosmos. His latest, The Grand Cosmic Story, is out now!
Metabolism, not cells or genetics, may have begun life on Earth was originally published in Starts With A Bang! on Medium, where people are continuing the conversation by highlighting and responding to this story.













{kind=link}